毛色関連遺伝子のDNA多型を
用いた豚の品種識別技術
三橋 忠由
◎最優秀賞:研究開発部門家畜ゲノム研究プログラムチーム遺伝子利用技術開発グループ |
はじめに
平成10年6月ごろ、DNAレベルでの豚の品種識別を目指し、毛色関連遺伝子のDNA多型を用いた豚の品種識別に関する研究を開始した。平成11年2月ごろ、本研究のなかから得られた、毛色に関連する2つの遺伝子の塩基配列情報を用いれば、バークシャー種を含むわが国で用いられる主要な豚品種について純粋種か、交雑種かを知ることができることが明らかとなり、当時問題になりはじめた「純粋黒豚」の識別に有用ではないかと考えるに至った。ここにいう「純粋黒豚」とは、単なる黒い色の豚のことではない。鼻端、尾端と4肢端の計6ヵ所が白く、「六白」といわれているバークシャー種のことである。バークシャー種の脂肪は融点が高く、肉には甘みがあり特においしいとされてきた。しかし、1腹産子数が少なく、体重の増加も遅いため飼養頭数はじょじょに減少し、平成9年2月時点では、全国で約12万頭(全体の約1.8%)となっていた。
最近、スーパーでは「黒豚」の表示を多くみかけるようになったが、消費者等から「本当の黒豚であるか疑問」「出荷量に比べて黒豚が氾濫しすぎている」、また「ランドレースなどと交配すると白い豚になる。黒豚が血統に入っていても、白い豚を黒豚と表示するのはどうか」といった意見があがっていた(畜産局)。
このため、農水省では「黒豚」についての意見を公募し、その定義(表示)を、純粋バークシャー種同士の交配による産子、とすることとした。ここで、食肉になった段階で純粋バークシャー種であることを、どのように識別するかが問題となっていたが、本研究課題である「毛色関連遺伝子のDNA多型を用いた豚の品種識別」が問題解決に貢献することとなった。
着目した遺伝子は、毛の色に関連するMC1R(メラノサイト刺激ホルモンレセプター:メラノコルチン1レセプター)遺伝子と、色素細胞等の増殖に関連するとされるKIT遺伝子の2つである。理由は後述するが、1つの遺伝子の配列型だけでは主要な品種を区別することはできない。2つの遺伝子の情報を用いることによってはじめて、ランドレースおよび大ヨークシャー種(白色)、デュロック種(褐色)、ハンプシャー種(白黒斑)、バークシャー種(白黒斑)、メイシャン豚(中国黒色種)の食肉試料での識別が可能になった(当稿では以後「種」を省略して記述する)。
1.毛色関連遺伝子について
ブタの毛色は、家畜化の過程で人為的選択圧を強く受けてきた形質であり、野生型(アグーチ;Agouti)からさまざまな色が出現してきた。黒(Black)、ドミノ斑(Domino spotting)、赤(Red)、白ベルト(White belt)は、中近東と地中海地方起源であり、優性白色(Dominant white)は、北ヨーロッパ起源と考えられている。現在までの長年にわたる交配実験の結果、ブタの毛色多型に関する遺伝子座は八つ、すなわちA(Agouti)、C(Albinism)、D(Dilution)、E(Extention)、He(Hereford)、I(Dominant white)、Be(White Belt)、R(Red-eye)が推定され、そのうちの5つ(A、E、He、I、Be)には毛色多型に関与する遺伝子の存在が推定されている。さらに近年、毛色の多型についての分子遺伝学的研究がマウスとヒトを中心に進展し、関与する遺伝子の知見が増大した。現在、わが国でおもに用いられているブタ品種も、特徴ある毛色、あるいは毛色パターンを示している。すなわち、ランドレース種と大ヨークシャー種は白色、デュロック種は赤(褐色)、バークシャー種は黒地に鼻先と尾、並びに足先が白い六白といわれる品種であり、毛色や体表面の紋様が品種を示す特徴の1つとなっている。
「品種は通常、毛の色と体表面の紋様パターンによって識別されている」、ということは「毛色および紋様を支配する遺伝子には、明らかな品種間の違いが存在するであろう」と思われた。このような考えのもとに、毛色および紋様に関連するとされる2種類の遺伝子、メラニン細胞刺激ホルモンレセプター(MC1R)遺伝子およびKIT遺伝子の多型を解析対象として、各ブタ品種に特徴的な配列を明らかにすることを試みた。
2.遺伝子塩基配列の測定
2つの遺伝子について、品種間の塩基配列の違いは特定領域をPCR増幅して、DNAシーケンサーにより直接に塩基配列を決定することによりみいだした。その後、塩基配列の違いが明らかになった部分を、簡易に制限酵素断片長多型によって検出することを試みた。制限酵素断片長多型はRFLP(Restriction fragment length polymorphisms)と呼ばれ、生物の個体間で、遺伝子の塩基配列の異なる部位に制限酵素切断部位が存在すると、制限酵素切断断片のサイズが異なることを利用して、その遺伝子の多型を解析するものである。3.MC1R遺伝子
MC1R遺伝子はメラニン細胞において、メラニン合成を促進するメラニン細胞刺激ホルモンのレセプター遺伝子であり、ブタ第6染色体の短腕にある。今回、ランドレース(白色)、大ヨークシャー(白色)、デュロック(褐色)、バークシャー(黒色)、ハンプシャー(黒色)、メイシャン(黒色)、モンカイ(黒色)、ニホンイノシシ(野生褐色)について、MC1R遺伝子の第二膜貫通領域を含む664bpの塩基配列を調べた。
各ブタ品種のMC1R遺伝子の塩基配列決定を行った結果、八ヵ所に塩基配列の違いを認めた。違いの認められた部分をアミノ酸の翻訳単位コドンで示した(表1)。コドン90、97、116、119、159、161、163、238において塩基置換が観察され、コドン116、163はアミノ酸の変化を伴わない置換、その他の変異はすべてアミノ酸の変化を伴う置換であった。なお、コドン番号はマウスのMC1R遺伝子の配列の番号に倣った。
表1のなかの配列番号M1はニホンイノシシ六個体からの配列である。ニホンイノシシ二個体はM1およびM2のヘテロ型であった。M3に示す塩基配列は、大ヨークシャー6個体、ランドレース2個体、バークシャー8個体、ハンプシャー8個体からのものである。また、ランドレースでは上記のM3に示す塩基配列のホモ型2個体以外に、M3およびM4のヘテロ型が2個体、M4のホモ型が2個体であった。M5はデュロック6個体からの配列である。メイシャン6個体とモンカイ3個体を調べた結果、すべての個体はM6に示すタイプのホモ型であった。
4.MC1R遺伝子の簡易な多型検出
4つの制限酵素(HhaI、MaeIIあるいはNLaIII RcaI)を用いた切断パターンの組合わせによりタイプ分けを行った。すべてのブタ品種において、塩基配列から予想されたとおりのRFLPの結果を得た。すなわち、| (1) | 制限酵素HhaIで消化した場合、デュロック対ランドレース、大ヨークシャー、メイシャン、バークシャー、ハンプシャーの識別が可能であった。 | |
| (2) | 制限酵素MaeIIあるいはNIaIIIで消化した場合、メイシャン対ランドレース、大ヨークシャー、デュロック、バークシャー、ハンプシャーが識別可能であった。 | |
| (3) | 制限酵素RcaIで消化した場合、デュロック、メイシャン対ランドレース、大ヨークシャー、バークシャー、ハンプシャーの識別が可能であった。 |
しかし、西洋白色種(ランドレース、大ヨークシャー)とバークシャー、ハンプシャーは塩基配列が同一のため、ここで取上げたMC1R遺伝子の多型からは、西洋白色種とバークシャーを識別することはできなかった。
5.KIT遺伝子
MC1R遺伝子の情報からだけでは、最も重要となる西洋白色種(ランドレース、大ヨークシャー)対バークシャーの識別ができなかった。そこで、毛色に関連するとされるもう1つの遺伝子、KIT遺伝子の塩基配列調査を行った。KIT遺伝子は第8染色体の短腕に存在し、ヒトでは約7万個のDNA分子から構成されている。 KIT遺伝子のエクソン16から19にわたる約4Kbpについて、上記の八品種に加えヨーロッパイノシシ(野生褐色)とユカタンマイクロブタ(黒色)の塩基配列を調べた。品種間で配列の違いがみいだされた部分を抜粋して示した(表2)。
大ヨークシャーおよびランドレースが持つ優性白色の理由が、c-KIT遺伝子の重複と重複遺伝子における変異によることがJohansson等により示唆されている。すなわち、重複した一方の遺伝子にはイントロン18に四塩基対の欠失がある。さらにエクソン17とイントロン17の境界の変異、すなわち、イントロン17の最初の塩基がグアニン(G)からアデニン(A)に変化することにより、その転写産物はエクソン17を欠く。このことについて、われわれの研究のなかでも追試し、すべての試料において矛盾のないことが確認された。すなわち、西洋白色種と有色種の違い(表2の1313番目)、イントロン18内の4塩基対の有無(表2の3884〜3887番目)が確認された。
この塩基配列情報により、西洋白色種と有色種が識別可能となり、前述のMC1R遺伝子の情報と組合わせ、主要な品種識別は可能になった。しかし、同じ白黒斑を持つバークシャーとハンプシャーの識別はまだ可能ではなく、後述するイントロンの塩基配列情報を用いる必要があった。
6.バークシャーとハンプシャーの識別
バークシャーとハンプシャーはどちらも白黒の体表面紋様を持ち、前述までの配列情報では識別できなかった。だが、KIT遺伝子のイントロン部分に両品種の識別に用いることができる多型が存在していた。イントロン部分の配列比較の結果、以下のような塩基置換の特徴が認められた。| (1) | バークシャーの配列はニホンイノシシ、メイシャン、モンカイの配列と類似しており、東洋起源と推察される。 | |
| (2) | 一方、ハンプシャーはヨーロッパイノシシ、大ヨークシャー、ランドレース、デュロックの1部と類似しており、西洋起源と推察される。 | |
| (3) | また、鹿児島県産の1部のバークシャーの配列はユカタンマイクロブタと類似していた(起源未定とした。)。 |
このようなイントロンの塩基配列の解析から、バークシャーは東洋起源と考えられる配列を持ち、KIT遺伝子959番目の塩基多型により識別できることが明らかになった。また、ハンプシャーは西洋起源と考えられる配列を持つことが明らかになり、KIT遺伝子2600番目の塩基多型により識別できることが明らかになった。
以上のように、本研究で用いた2つの毛色関連遺伝子の塩基配列多型を検出することにより、日本における主要なブタ品種、白色品種(ランドレース、大ヨークシャー)、デュロック、バークシャー、ハンプシャーを生肉あるいは加工肉からでも識別可能なことが明らかとなった(図1)(表3)。
表3 5種類の制限酵素を用いたハム・ソーセージ等加工肉における遺伝子診断の結果例
図1 毛色に関与する2つの遺伝子の多型によるバークシャー種の識別
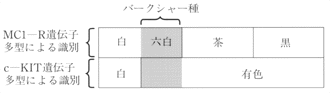
おわりに
本研究で用いた2つの毛色関連遺伝子の塩基配列多型を検出することにより、日本における主要なブタ品種、白色品種(ランドレース、大ヨークシャー)、デュロック、バークシャー、ハンプシャーを生肉あるいは加工肉からでも識別可能なことが明らかとなった。 遺伝子を用いた識別では、同じ遺伝子型を保持していれば同一の品種と識別される。すなわち、本方法は検査に供する加工肉あるいは生肉試料が前述の2つの遺伝子について、純粋バークシャー種と同じ遺伝子型を持つかどうかを検出するものである。
〈用語説明〉
| (12) | Kbp(キロベースペアー) ベースペアーとは塩基(DNA)対のこと。何対の塩基対で構成されているかを示すときに用い、遺伝子が大きい場合にはK(キロ)単位(1Kbp=1000塩基対)で示す。 |
(筆者は農水省畜産試験場育種部・遺伝子機能研究室長)
遺伝子の部分でアミノ酸に翻訳される部分をエクソン、翻訳されない部分をイントロンと呼ぶ。
塩基配列のタイプのこと。
塩基配列のうち一定の配列型のみを認識し、当該部分を切断する酵素のこと。
個体の遺伝子は両親から受継いでいる。父親と母親から受継いだ遺伝子型が同じ場合はホモ型、異なる場合はヘテロ型と呼ぶ。
塩基配列のうち個体によって配列が異なる(入替わっている)部分のこと。
遺伝子の塩基配列は3つの塩基で1つのアミノ酸に翻訳される。この塩基3つの単位をコドンという。
細胞表面にあるレセプターには細胞内部に刺激を伝えるために細胞膜を貫通して存在している。細胞膜にも無論厚さが存在し、レセプタータンパクの1部は膜と重なっている。この領域が膜貫通領域と呼ばれる。MC1Rレセプターは七つの膜貫通領域を持つ。
細胞表面、時には細胞内にあり、ホルモン等に結合し、ホルモンの刺激を他の生体反応へ伝える働きを持つ。
DNAの配列(塩基配列)が個体間で異なること。すなわち、1つの型でなく多くの型を持つこと。
白血病(癌)ウイルスと類似の配列を持つ遺伝子として発見された。癌遺伝子と同様、細胞の増殖に関連する。色素細胞の増殖に関連すると考えられ、体表面の黒白斑の形成に関与することも考えられる。ブタでは第8染色体上にある。
髪の毛の色、皮膚の色の素であるメラニン色素を合成する色素合成細胞(メラノサイト)細胞の表面にある蛋白質であり、色素合成のホルモン刺激を受取り、細胞内部に色素合成の信号を伝える。このMC1R蛋白質を作るもとになるのがMC1R遺伝子である。ブタでは第6染色体上にある。